
Ecologists are frequently challenged with the task of making formal predictions about how populations will respond to different forms of management or perturbations. Whether making predictions about optimal harvest policies, invasive species control, or population recovery, the same principles apply. The broadly applicable key determinants of how a population responds to mortality events are (i) the general life-history strategy and (ii) how density dependent processes regulate the population dynamics rates.
The role of life history
General life-history strategy refers to the fact that there is more than one way to persist as a species. The most basic way to think about this is using the model of r and K selection theory. Life-history strategies that are r or K selected represent two “book ends” of a gradient of possible strategies. Being r selected means that the population has a high potential for growth. In other words, individuals of that species have a large ![r-k-selection[8]](https://dgwinn.files.wordpress.com/2017/12/r-k-selection8.jpg?w=436&h=270) reproductive capacity, giving the population its high potential for rapid growth. These species tend to grow fast, mature early and have short life spans. They will be able to sustain high exploitation rates, will be difficult to control with removal efforts, and will recover quickly from mortality events.Alternatively, being K selected means that the population does not have a high potential for growth but relies on large population sizes to persist (I pinched the above figure from link). These species tend to grow slow, mature late, and have long life spans. They cannot sustain high exploitation, will be more responsive to control via removal, and will require longer time periods to recover from mortality events.
reproductive capacity, giving the population its high potential for rapid growth. These species tend to grow fast, mature early and have short life spans. They will be able to sustain high exploitation rates, will be difficult to control with removal efforts, and will recover quickly from mortality events.Alternatively, being K selected means that the population does not have a high potential for growth but relies on large population sizes to persist (I pinched the above figure from link). These species tend to grow slow, mature late, and have long life spans. They cannot sustain high exploitation, will be more responsive to control via removal, and will require longer time periods to recover from mortality events.
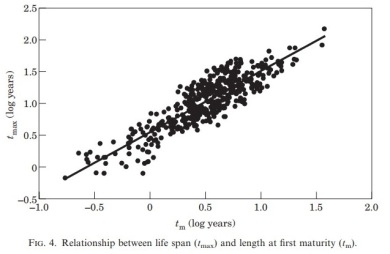
Using general life-history strategies to predict population response to mortality events tends to be the easier part of the puzzle. This is because it is fairly easy to make predictions with a population model if you know some of the basic aspects of a species life history such as, life span and age at maturation. Furthermore, it’s pretty easy to get information on a species life history or take good guesses based on published life-history invariants. For  example the mean age that a fish species matures is highly related to its mean total life span (Froese and Binohlan 2000, Fig. above), which is a function of its natural mortality rate (Hoenig 1983, Fig. to left), which is strongly related to its growth rate (Jensen 1996). The difficult part tends to be understanding how density dependent processes influence the population dynamics rates.
example the mean age that a fish species matures is highly related to its mean total life span (Froese and Binohlan 2000, Fig. above), which is a function of its natural mortality rate (Hoenig 1983, Fig. to left), which is strongly related to its growth rate (Jensen 1996). The difficult part tends to be understanding how density dependent processes influence the population dynamics rates.
The role of density dependence
Life-history, in a sense determines the maximum rate at which a population can grow, whereas density-dependent processes regulate that growth rate. For example, the biology of a species may determine its maximum per capita birth rate, but the proportion of juveniles surviving to adulthood may be related in part to the density of those juveniles relative to resource availability. One interesting model of this phenomenon is Foraging Arena Theory (Ahrens et al. 2012). The idea of Foreging Arena Theory was presented as a mechanism for density dependent compensation in juvenile fish survival first by Walters and Juanes (1993) and has more recently been reviewed by Ahrens et al. (2012). The theory describes juvenile fish as existing in two states, one invulnerable to predation and the other vulnerable to predation, and the proportion of time spent in either state as a function of density. The basic idea is that juvenile fish spend part of their time in refuges from predation such as near structure or in schools. In these refuges, the local  resources will be depleted; thus, fish will leave the refuge to enter “foraging arenas” to feed where their predation risk is increased. The greater the densities of juvenile fish in the refuges, the greater the depletion of local resources, and the greater the need to risk
resources will be depleted; thus, fish will leave the refuge to enter “foraging arenas” to feed where their predation risk is increased. The greater the densities of juvenile fish in the refuges, the greater the depletion of local resources, and the greater the need to risk  predation by entering into the foraging arenas. Thus, higher densities leads to less per capita resources, increased engagement in risky foraging behavior, and decreased mean survival of juvenile fish…..i.e., density dependent survival. But how does this impact the way a population responds to mortality events? For populations where the gradient in predation risk between the refuge
predation by entering into the foraging arenas. Thus, higher densities leads to less per capita resources, increased engagement in risky foraging behavior, and decreased mean survival of juvenile fish…..i.e., density dependent survival. But how does this impact the way a population responds to mortality events? For populations where the gradient in predation risk between the refuge
Higher densities leads to less per capita resources, increased engagement in risky foraging behavior, and decreased mean survival of juvenile fish.
and the foraging arena is great, you can expect large increases in survival when high densities are reduced. This is what is referred to as “density dependent compensation” and is a form of resilience because the total juvenile fish surviving to adulthood may remain constant even when densities of juveniles (or breeding adults) are reduced. Alternatively, for populations where the gradient in predation risk between the refuge and the foraging arena is small, you can expect little change in mean survival across juvenile fish densities, thus, low levels of density-dependent compensation and less resilience to mortality events.
The example of redfin perch control in Australia
An example of how life-history and density-dependent processes come together to determine optimal management strategies is the case of redfin perch control in Australia. Redfin perch were introduced into Australia in the late-1800s to develop a recreational fishery (Weatherley 1977). Today, redfin perch are considered an invasive pest in Australia because they prey upon native fish species causing a conservation concern (Barrett et al. 2014, Lintermans et al. 2014, Beatty and Morgan 2016). Recently, in Lake Purrumbete, Victoria there has been motivation to attempt some form of control of the redfin perch population (Ingram 2016). This system is fairly unique because it is managed mainly for recreational fishing with stocking programs for multiple salmonid species including rainbow trout, brown trout and Chinook salmon. Redfin perch densities are high in the lake with evidence of stunting. The motivation for  controlling redfin perch is driven mainly by the recreational fishery because large numbers of small redfin perch are caught in exclusion of more desirable species resulting in poor fishing quality. There is also a belief by resource users that redfin perch are out competing other salmonid species for resources resulting in poor fish condition. However, the control of redfin perch may be difficult due to its life history and density-dependent regulation.
controlling redfin perch is driven mainly by the recreational fishery because large numbers of small redfin perch are caught in exclusion of more desirable species resulting in poor fishing quality. There is also a belief by resource users that redfin perch are out competing other salmonid species for resources resulting in poor fish condition. However, the control of redfin perch may be difficult due to its life history and density-dependent regulation.
So, what aspects of the Lake Purrumbete redfin perch population would lead you to conclude that population control may be difficult? Firstly, redfin perch are most definitely an r selected species with short generation times and a very high population growth potential. The characteristics that support this conclusion are their small maximum length (~400 mm), their short average life span (~13 years), their high natural mortality rate (~0.34), their tendency for early maturation (~1-2 years). These characteristics alone indicate that the species probably has the potential for fast population growth.
In addition to the r selected life history, redfin perch demonstrate a variety of density-dependent regulatory mechanisms. For example, larval redfin perch are closely associated with vegetation in the shallow littoral zone of water bodies. This behavior of larval fish can result in a high gradient between the predation risk within this refuge and the predation risk in the foraging arena of more exposed water. Thus, we expect high levels of density-dependent compensation in juvenile survival simply due to competition for resources in the refuge and the increased predation risk outside of the refuge. In addition to this, adult redfin perch are known to be cannibalistic on juveniles.  This inter-cohort interaction can greatly enhance density-dependent larval survival because a reduction in adults not only leads to fewer larvae (less competition), but also results in less predation (in the form of cannibalism). Populations with cannibalistic behavior such as this can demonstrate increased numbers of juveniles surviving to adulthood when adult abundance is reduced. This phenomenon is often termed “over compensation” (Zipkin et al. 2009).
This inter-cohort interaction can greatly enhance density-dependent larval survival because a reduction in adults not only leads to fewer larvae (less competition), but also results in less predation (in the form of cannibalism). Populations with cannibalistic behavior such as this can demonstrate increased numbers of juveniles surviving to adulthood when adult abundance is reduced. This phenomenon is often termed “over compensation” (Zipkin et al. 2009).
Another form of density-dependent compensation common in redfin perch populations is through adult growth rates. Redfin perch can demonstrate stunted growth when adult population densities are high. Thus, any reduction in densities can result in faster adult growth rates, increased per capita fecundity, and the potential for increased cannibalism on juveniles. You can see how it can be quite difficult to synthesize these different mechanisms of density dependence to produce a predicted response to mortality in your head. The solution to this, of course, is to use a population model that accounts for these sources of density dependence to simulate and create formal predictions about the response of redfin to mortality events.
In a recent paper, a colleague and I performed a simulation study to evaluate different removal options for redfin perch in Lake Purrumbete (Gwinn and Ingram 2018). The modeling indicated that any exploitation of redfin will likely result in increased recruitment unless the exploitation is high enough to overcome the species ability to compensate. This condition is frequently termed recruitment overfishing and for redfin perch would only occur when exploitation rates are very high (perhaps unrealistically high). The other condition was when exploitation targeted fish that were smaller than the size when they start to engage in cannibalism. Targeting small fish directly reduced the number of small fish, reduced densities of fish recruiting to adulthood, thus increasing growth rates, and maintaining the large adult fish that regulate recruitment through cannibalism. The predicted effect was a transition of the redfin perch population from high densities of small fish to moderate densities of large fish, which may be more desirable to anglers.
References
Gwinn, D.C., and Ingram, B. (2018) Optimizing fishery characteristics through control of an invasive species: strategies for redfin perch control in Lake Purrumbete, Australia. Marine and Freshwater Research https://doi.org/10.1071/MF17326
Gwinn, D.C., and Allen, M.S. (2010) Exploring population-level effects of fishery closures during spawning: an example using largemouth bass. Transactions of the American Fisheries Society 139, 626-634.
Allen, M.S., Rogers, M.W., Catalano, M.J., Gwinn, D.C., and Walsh, S.J. (2013) Assessing the Potential for the Angling of Nesting Males to Influence Largemouth Bass Recruitment Should Consider the Quality of Nesting Individuals: Response to Comment. Transactions of the American Fisheries Society 142, 419-421.
Allen, M.S., Rogers, M.W., Catalano, M.J., Gwinn, D.C., and Walsh, S.J. (2011) Evaluating the Potential for Stock Size to Limit Recruitment in Largemouth Bass. Transactions of the American Fisheries Society 140, 1093-1100.
Shaw. S.L., and Allen, M.S. (2016) Quantifying Recruitment Compensation in Florida Largemouth Bass, with Implications for Fisheries, Transactions of the American Fisheries Society 145, 462-475.
Ahrens, R.N.M., Walters, C.J., and Christensen, V. (2012) Foraging arena theory. Fish and Fisheries 13, 41-59.
Froese, R., and Binohlan, C. (2000) Empirical relationships to estimate asymptotic length, length at first maturity and length at maximum yield per recruit in fishes, with a simple method to evaluate length frequency data. Journal of Fish Biology 56, 758-773.
Walters, C.J., and Juanes, F. (1993) Recruitment Limitation as a Consequence of Natural Selection for Use of Restricted Feeding Habitats and Predation Risk Taking by Juvenile Fishes. Canadian Journal of Fisheries and Aquatic Science 50, 2058-2070.
Hoenig, J.M. (1983) Empirical use of longevity data to estimate mortality rates. Fishery Bulletin 82, 298-903.
Jensen, A.L. (1996) Beverton and Holt life history invariants result from optimal trade-off of reproduction and survival. Canadian Journal of Fisheries and Aquatic Sciences 53, 820–822.
Ingram, B.A. (2016) Options for managing redfin perch in Lake Purrumbete to improve recreational fishing. Recreation Fishing Grants Program Research Report.

Nice blog post Dan. Thanks for writing.